
PIE-LTER Research Objectives and Project Summary
Human activities in rivers and watersheds have altered enormously the
timing, magnitude and nature of inputs of materials such as water, sediments,
nutrients and organic matter to estuaries (Hopkinson and Vallino 1995). One of
the dominant themes of estuarine research in recent decades has been the
effects of inorganic nutrients on eutrophication of coastal waters. Studies
have detailed how inorganic nutrient inputs from land influence primary
productivity, the depletion of oxygen, habitats, and trophic relationships in
coastal waters. However, another important but neglected linkage between land
and coastal waters is the input of dissolved and particulate organic carbon and
organic nitrogen. Organic nitrogen inputs are frequently greater than inorganic
nitrogen inputs and in many cases the input of organic carbon from land is
greater than the in situ production of organic carbon. Despite the
quantitative importance of organic matter inputs from land, there are few data
on its actual use in coastal waters and even fewer on effects of organic matter
loading on coastal ecosystem trophic structure and trophic productivity. Yet,
in other ecosystems such as rivers, lakes, and the deep-sea, effects of
allochthonous organic matter inputs on trophic structure and ecosystem metabolism
have been well documented.
Despite an awareness of large scale, long-term changes in watersheds, we do not fully understand the consequences of activities such as damming of rivers, land use conversion, and removal of floodplains. Often human influences on river systems are opposing. For instance land clearing increases drainage basin sediment yield while damming decreases sediment discharge.
Climate variability and long-term patterns of climate change also can have immense effects on the timing, magnitude and nature of material inputs. Infrequent storms can accomplish in days what normally occurs over decades. For instance, heavy rainfall associated with tropical storm Agnes increased Susquehanna River flow to record levels, which resulted in 30 years of "normal" sediment discharge occurring in one week (Hirshberg and Schubel 1979).
Variations and long-term increases in sea level affect estuaries from their seaward end. Tides and sea level have significant effects on water and marsh sediment salinity, plant community composition (Olff et al. 1988), primary production (Morris and Haskin 1990, Morris 1995), access to marsh surface habitats by nekton (Rozas 1995) and sediment accretion (Callaway et al. 1996).
The interaction of organic matter and nutrient inputs from land and variations in the external forcings (climate, land use, river discharge, sea level) regulating estuarine mixing and residence time will dictate the extent of nutrient and organic matter processing during estuarine transport and will determine the spatial patterns of productivity and trophic structure. The PIE-LTER research program focuses on the following question and hypotheses:
How will trophic structure and primary and secondary productivity in estuaries be affected by changes in organic matter, nutrient and water fluxes caused by changing land cover, climate and sea level?
Hypothesis 1. The interaction of inorganic nutrients with the quantity and quality of organic carbon and organic nitrogen plays an important role in determining the trophic structure, production and efficiency of estuarine food webs.
Hypothesis 2. The variability in land, ocean and atmospheric forcing is a key component determining the fate of allochthonous and autochthonous materials and the location and magnitude of primary and secondary production.
We are testing these hypotheses through 1) short- and long-term measurements of the fluxes of dissolved and particulate organic carbon and organic nitrogen entering estuaries from land, marshes and the ocean, 2) short and long-term experiments that determine the effects of various nutrient and organic matter inputs and interactions on the flow of C and N through pelagic and benthic food webs, and 3) simulation models that examine the effects of land use changes on food web transformations. Our research integrates estuarine biogeochemistry with studies of food webs and population biology of higher trophic levels.
The study consists of 1) site-specific research at Plum Island Sound, and 2)
comparative ecosystem research, especially at North Inlet, SC and Wells, ME
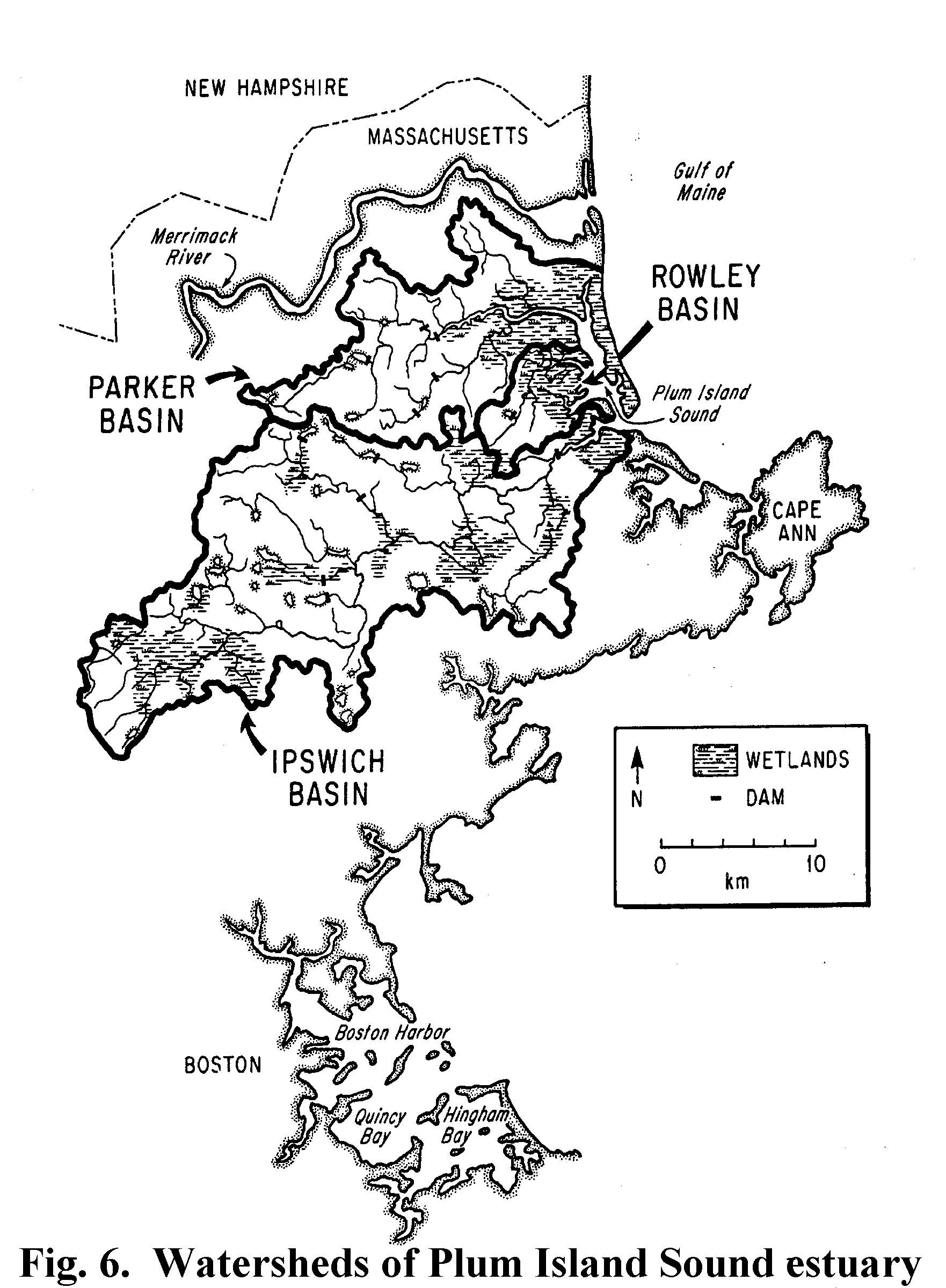
national estuarine research reserves. Plum Island Sound is in the cold-water
Acadian Biogeographic Province (Fig 6) and lies at the interface of a
thin soiled, formerly glaciated New England watersheds and the highly
productive Gulf of Maine. Three watersheds with differing levels and rates of
development drain into the Plum Island Sound tidal marsh estuary providing a
wide range in the quantity and quality of inorganic and organic matter loading.
In addition to experimental manipulations, our research program uses natural
experiments by taking advantage of these watershed differences within and
between estuaries to determine the importance of inorganic and organic matter
interactions in controlling trophic structure, trophic production and trophic
efficiency.
Hypothesis 1. Organic Matter - Nutrient Interactions and
Trophic Structure:
The input of organic matter from land to coastal waters is substantial. Over
the entire earth, 1-2% of terrestrial NPP (net primary production) or 1-14 gC m-2yr-1
(Meybeck 1982) is exported to estuaries; this is equivalent to 28% of estuarine
NPP (Wollast and Billen 1981). The majority of organic carbon is exported as
dissolved organic carbon (DOC); the remainder is particulate organic carbon
(POC). Organic nitrogen is also exported primarily in the dissolved form and at
the global level riverine export of organic N surpasses that of dissolved
inorganic N (DIN)(Meybeck 1982); dissolved organic nitrogen (DON) in rivers
comprises 68% of the total dissolved nitrogen (TDN). In the Plum Island System,
DON and DOC comprise 60-80% and 92-96% of TDN and TOC, respectively.
Another major source of organic matter in estuaries is intertidal marshes. In marsh-dominated systems along the east and Gulf coasts of the U.S., marsh inputs of organic matter to adjacent tidal creeks often greatly exceed in situ production (Hopkinson 1988).
Estuaries are not merely conduits transporting organic material from rivers to the sea. Estuarine biota use this organic matter, often creating a region of heterotrophy, where respiration exceeds local primary production (Smith and Mackenzie 1987, Hopkinson and Vallino 1995). Most of this respiration is by bacteria (Hopkinson et al. 1989). When supported by terrestrial or marsh organic matter with high C/N ratios, estuarine bacteria compete with primary producers for nutrients (Goldman et al. 1987a,b, Vallino et al. 1996, Wheeler and Kirchman 1986, Kirchman et al. 1989, Tupas and Koike 1990, Toolan et al. 1991, Currie and Kalff 1984). If all the terrestrial organic input was decomposed by bacteria with a 40% growth efficiency, they would use twice the global flux of inorganic N from the land. While much of this organic matter is refractory, a great deal is processed in the estuary and we need to determine its effects on estuarine trophic structure and efficiency.
The delivery of organic matter and nutrients has a major effect on estuarine food webs. For example, the numbers and activity of the flagellates and other members of the microbial loop increased over a eutrophication gradient in Narragansett Bay waters (Hobbie and Cole 1984) and in a freshwater mesocosm (Hobbie and Helfrich 1988). Both zooplankton and benthic animals increased in numbers in response to sewage sludge in a MERL experiment (Oviatt et al. 1987). Several studies have correlated fish production with river discharge and the delivery of organic matter to estuaries (e.g., Sutcliffe 1972, Armstrong 1982). Fish production is generally low (per unit C input) when organic matter loading is high and nutrient availability is low, presumably because of food web inefficiencies (Quiros and Baigun 1985, Welcomme 1985). Blackwater rivers, rich in DOM (usually humics) but low in nutrients, typically have low fish production (Welcomme 1985). Information on the processes underlying these correlations is largely lacking.
The few studies discussed above deal solely with organic carbon; the dearth of data is even greater for organic nitrogen. For example, even the careful budget of Nixon and Pilson (1984) for nitrogen in Narragansett Bay omits the input of organic nitrogen. Studies of DON use in estuaries are generally limited to uptake measurements of labeled amino acids (e.g., Crawford et al. 1973) but amino acids make up only a small proportion of the bulk DON. Only very recently has there been a realization that a significant fraction of riverine DON is labile (Uhlenhopp et al. 1995, Seitzinger and Sanders 1997,1999, Wiegner and Seitzinger 2001) and that it may contribute to the nutrition of phytoplankton (Bronk and Glibert 1993). There are no measures of incorporation of DON from land into higher trophic levels of the estuarine food web.
Hypothesis 2. Variability in Land, Ocean and Atmospheric Forcing:
All components of an estuarine ecosystem exhibit variation in space and time in
response to external forcings such as changes in climate, river discharge and
sea level or in response to internal, biogeochemical and biological processes
such as nutrient turnover, population cycles and fish migrations. By comparing
time scales of processes, the relative importance of various physical or
biological factors in controlling variation in process rates can be assessed
and limits placed on the ability of one process to affect another (Hatcher et
al. 1987). Processes that occur at similar scales are those likely to interact.
Our proposed research on the effect of changes in organic matter, nutrient and
water fluxes on estuarine trophic structure and production requires that we
evaluate variability in land and ocean forcings and assess the spatial and
temporal scales over which the effects of these changes are likely to operate.
Understanding the response of these systems to long-term changes in climate,
sea level and land cover first requires that we understand and can distinguish
short-term variation from long-term trends.
The important external forcings for the Plum Island Sound land margin ecosystem have different scales of variability. Precipitation averages 1.1 m yr-1 but ranges between 0.6 - 1.8 m yr-1. Day to day variability is considerably greater; during a major storm, rainfall can amount to 1/5 the annual mean. Over the past 70 years, precipitation has increased 2.9 mm yr-1 (Claessens et al. 2002). River runoff is more variable than rainfall. Daily discharge varies by a factor > 104 over the year and average annual discharge varies 5 fold. Material loading does not vary directly in proportion to discharge; DOC varies to a lesser degree (e.g., 1993-96 annual discharge varied 25% while DOC export varied 20%) while others vary more (e.g., 1993-96 POC export varied 220%). Tidal amplitude averages 2.9 m, but ranges from 2.6 -3.6 m during the neap-spring cycle with storm surges reaching 4.6 m. Sea level has been increasing 2.4 mm yr-1. Mean monthly sea level varies 15 cm over the year but has been as large as 30 cm. Mean annual sea level varies about 13 cm. Other forcings with considerable variability include insolation and temperature.
Estuarine water residence time is a metric that reflects much of the variability in external forcings. Average residence time of water originating in the upper Plum Island estuary decreases from 34 - 5 d as river flow increases from 0.01 to 10 m3 s-1 (Vallino and Hopkinson 1997). In the lower estuary these changes in flow have less effect, with residence time decreasing from 1.2 - 0.6 d. The influence of organic matter and nutrient inputs on the estuarine food web will be dictated by the time scales of important processes relative to water residence time. Examples of these processes include bacterial growth, organic matter decay, particulate organic matter settling, nutrient uptake and phytoplankton growth, and consumer growth relative. Benthic food webs will be less influenced directly by variations in residence time than pelagic webs but nevertheless will be indirectly influenced by changes in salinity and pelagic resources.
PIE-LTER research investigates the cause and effect relationships among the physical and biological processes that define estuarine structure and production. We must understand the basic interactions between physical and ecological processes before we can reliably predict the responses of estuaries to changes brought about by human activities (Geyer et al. 1997).
Synthesis - A Trophic Flow Model
We developed a trophic flow model that synthesizes our understanding of the
effects of organic matter and nutrient inputs on trophic structure and
function. A simplified diagram of the model illustrates how the major research
questions of the PIE LTER project fit together (Fig 7).
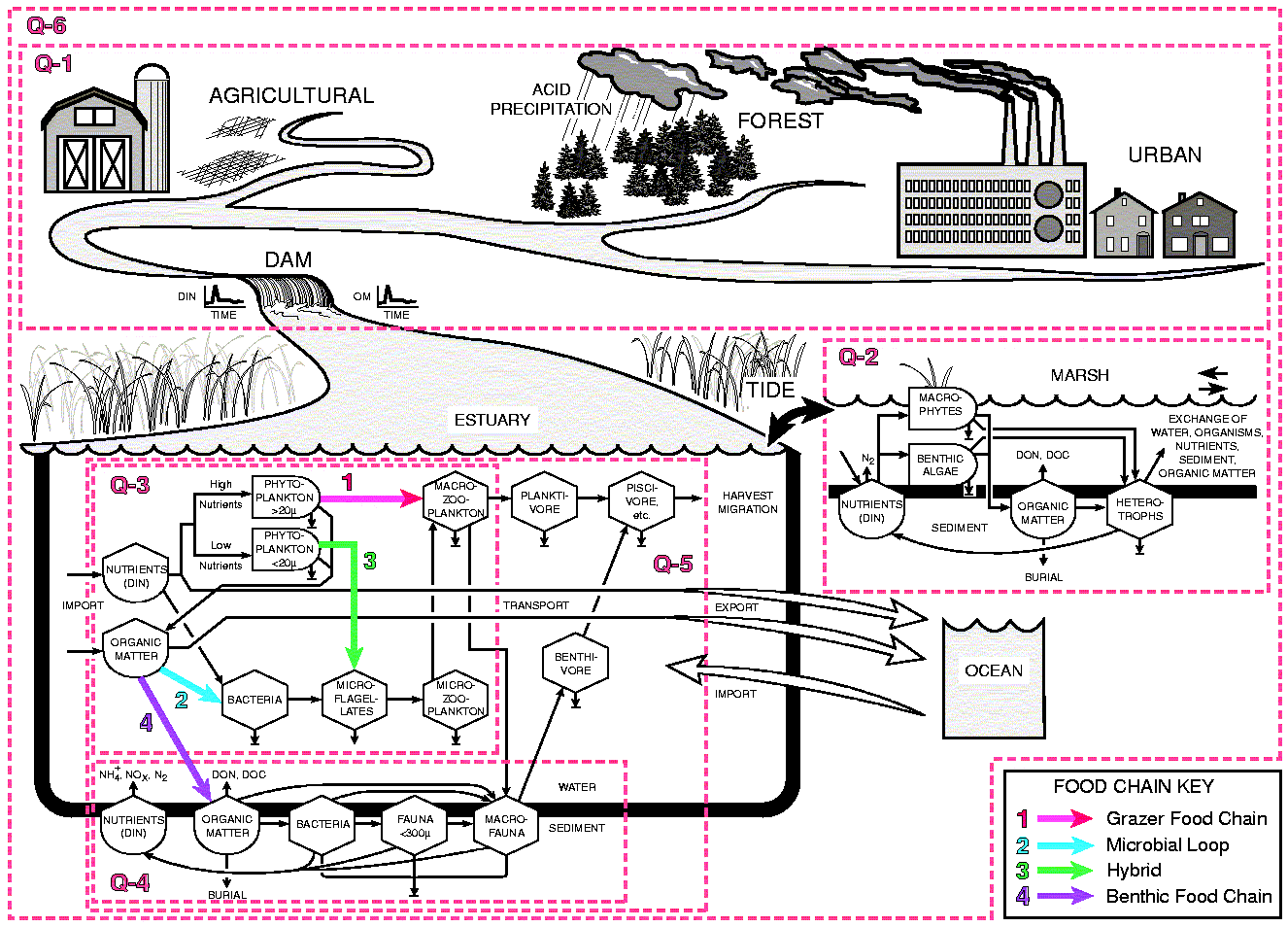
The model illustrates the possible interactions of inorganic nutrients with the quantity and quality of organic matter and their affect on trophic structure and trophic efficiency. Four major organic matter and nutrient pathways are portrayed: 1) the classical grazing food chain: inorganic nutrients - phytoplankton > 20 µm - macrozooplankton - planktivores - piscivores, 2) the microbial loop: organic matter - bacteria - microflagellates - microzooplankton - macrozooplankton - planktivores - piscivores, 3) a hybrid of the grazing food chain and the microbial loop: inorganic nutrients - phytoplankton < 20 µm - microflagellates - microzooplankton (the microbial loop) - macrozooplankton - planktivores - piscivores, and 4) a benthic food chain: organic matter - to sediments - bacteria - benthic meio - and macrofauna - benthivores - piscivores. The degree and rate at which organic matter flows through one pathway or another depends largely on the quality of the organic matter being decomposed and the availability of inorganic nutrients. High organic matter loading stimulates the microbial loop at the expense of phytoplankton while high nutrient loading stimulates the grazing food chain. We expect trophic efficiency to be partially controlled by organic matter quality and availability of inorganic nutrients for bacteria.
We have used our flow model to evaluate the effects of variations in organic matter and inorganic nutrient loading on trophic structure and production of higher trophic levels including piscivorous fish (Deegan et al. 1994). Development of the model and analysis of simulation results test the integrity of our understanding and help us guide research efforts. Two scenarios illustrate the effects of extremes in organic matter and nutrient loading:
Scenario 1. High Loading of Low Quality Organic Matter - Low DIN Loading: the major flow of carbon is through an inefficient microbial loop. Overall trophic efficiency is low; piscivore production is supported mostly by the benthos. The effect of the benthos is to increase the efficiency at which low quality organic matter is processed and transferred through the food web.
Scenario 2. Low Loading of High Quality Organic Matter - High DIN Loading: the major flow of carbon is through an efficient grazing food chain and secondarily through the benthos. Overall trophic efficiency is approximately twice that of scenario 1; piscivore production is supported by grazing and benthic chains. However, the benthos, by shunting more N out of the system via denitrification, causes a reduction in piscivore production.
Research - We focus LTER research around five primary research questions. The scope of each question and the manner in which each fits in the overall program is illustrated with numbered boxes around portions of our conceptual model (Fig 7).
Q1. What are the magnitude and temporal pattern of organic carbon and nitrogen and inorganic N loading from watersheds to the estuary?
Q2. How are tidal marsh processes and their connections to estuarine waters regulated by sea level, storms and water and material inputs from land and sea?
Q3. How do planktonic community structure and production respond to changes in organic matter, nutrients and water fluxes?
Q4. How do benthic use and recycling of nutrients and organic matter vary with changes in water fluxes and the quality and quantity of organic matter inputs?
Q5. How do the structure and function of higher
trophic levels respond to variations in organic matter, nutrients and water
fluxes?
![]()
