
PIE-LTER Research Objectives and Project Summary
Ecosystems at the land-sea interface play a major but poorly understood role in regional as well as global hydrologic, sediment, and biogeochemical cycles. Coastal ecosystems play a key role in the transformation, transport, burial and exchange of water and organic and inorganic carbon and nitrogen between land, atmosphere and the ocean. With an overwhelming majority of the human population living in the coastal zone and with runoff from entire continents funneling through estuaries and ocean margins, coastal systems are among the most heavily impacted ecosystems on the globe.
The biosphere is undergoing unprecedented change
as a result of human activities. Major global issues include
growth of the human population, land use change, climate change, altered
hydrologic cycles, and sea level rise. These are also important issues at
regional scales, as well as at the PIE LTER. Human population in the U.S.
continues to increase, especially in the coastal zone; we see similar dramatic
increases in population in the Ipswich and Parker River watersheds (Fig 1). 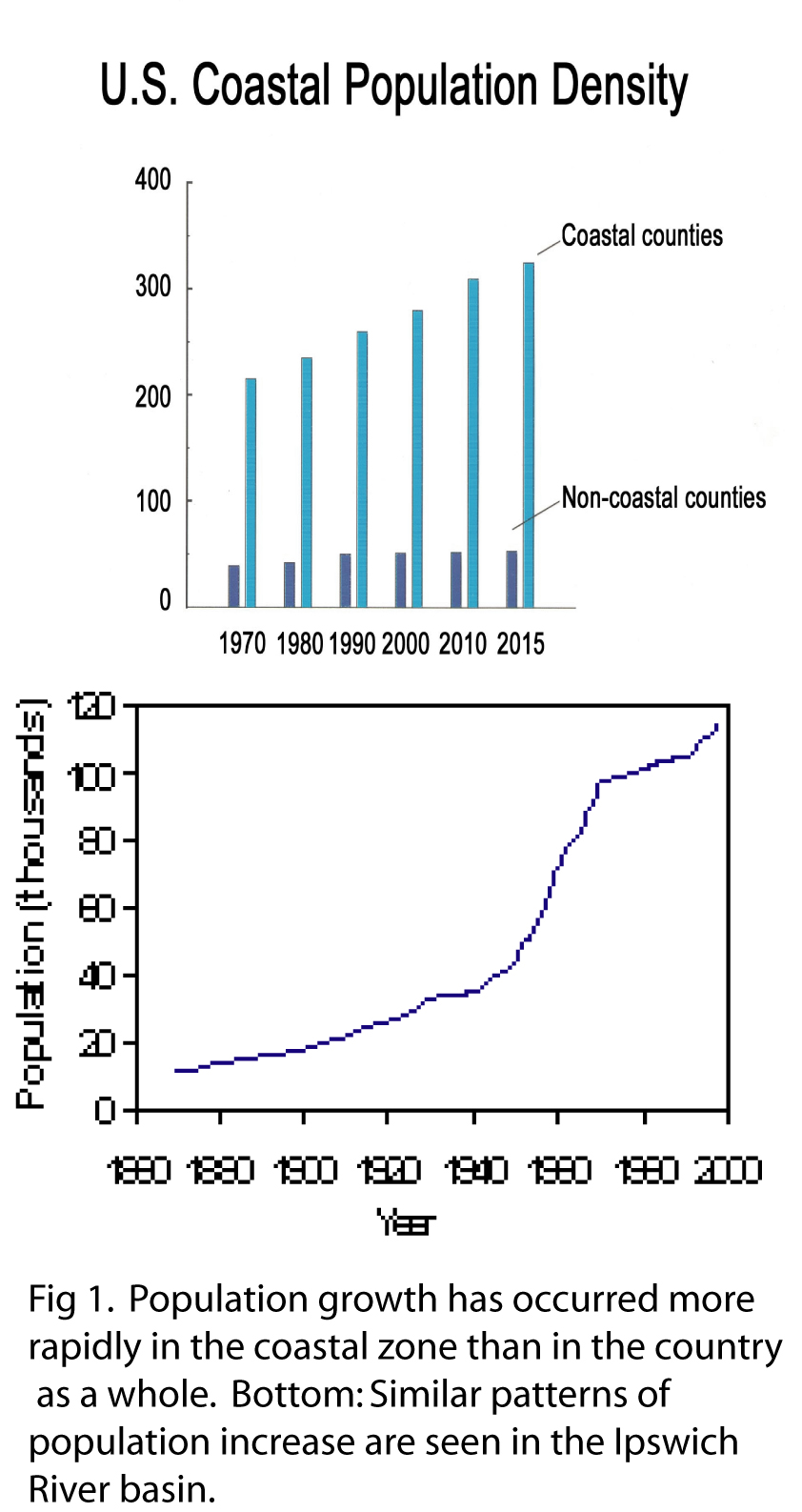 Land use has
changed substantially along the entire east coast of the U.S. over the past
century. The pattern of abandonment of agriculture, reforestation followed by
urbanization has occurred in the PIE watershed as well (Schneider and Pontius
2001) (Fig 2). There have been substantial changes in the
frequency of storms along the U.S. Atlantic coast, with the magnitude of change
in storminess increasing at more northern latitudes. At the PIE LTER
changes in century-long storm frequency are pronounced and precipitation and
evapotranspiration have increased significantly (Fig 3 Claessens
et al. 2006). In the future, we can expect to see more frequent, larger storms
and higher storm frequency variability as well. There have been decadal and
century-long changes in sea level at PIE as well (Fig 4). Therefore, the PIE LTER study can be
considered a microcosm for investigating the effects of these globally important
changes and it should thus be possible to transfer lessons learned from the PIE
LTER to other coastal regions. The biosphere is undergoing unprecedented
change as a result of human activities.
Major global issues include growth of the human population, land use
change, climate change, altered hydrologic cycles, and sea level rise. These are
also important issues at regional scales, as well as at the PIE LTER. Human
population in the U.S. continues to increase, especially in the coastal zone; we
see similar dramatic increases in population in the Ipswich and Parker
River watersheds (Fig 1). Land use
has changed substantially along the entire east coast of the U.S. over the past
century. The pattern of abandonment of agriculture, reforestation followed by
urbanization has occurred in the PIE watershed as well (Schneider and Pontius
2001) (Fig 2). There have been
substantial changes in the frequency of storms along the U.S. Atlantic coast,
with the magnitude of change in storminess increasing at more northern
latitudes. At the PIE LTER changes in century-long storm frequency are
pronounced and precipitation and evapotranspiration have increased significantly
(Fig 3 Claessens et al. 2006). In the future, we can expect to see more
frequent, larger storms and higher storm frequency variability as well. There
have been decadal and century-long changes in sea level at PIE as well
(Fig 4). Therefore, the PIE LTER
study can be considered a microcosm for investigating the effects of these
globally important changes and it should thus be possible to transfer lessons
learned from the PIE LTER to
other coastal regions.
Land use has
changed substantially along the entire east coast of the U.S. over the past
century. The pattern of abandonment of agriculture, reforestation followed by
urbanization has occurred in the PIE watershed as well (Schneider and Pontius
2001) (Fig 2). There have been substantial changes in the
frequency of storms along the U.S. Atlantic coast, with the magnitude of change
in storminess increasing at more northern latitudes. At the PIE LTER
changes in century-long storm frequency are pronounced and precipitation and
evapotranspiration have increased significantly (Fig 3 Claessens
et al. 2006). In the future, we can expect to see more frequent, larger storms
and higher storm frequency variability as well. There have been decadal and
century-long changes in sea level at PIE as well (Fig 4). Therefore, the PIE LTER study can be
considered a microcosm for investigating the effects of these globally important
changes and it should thus be possible to transfer lessons learned from the PIE
LTER to other coastal regions. The biosphere is undergoing unprecedented
change as a result of human activities.
Major global issues include growth of the human population, land use
change, climate change, altered hydrologic cycles, and sea level rise. These are
also important issues at regional scales, as well as at the PIE LTER. Human
population in the U.S. continues to increase, especially in the coastal zone; we
see similar dramatic increases in population in the Ipswich and Parker
River watersheds (Fig 1). Land use
has changed substantially along the entire east coast of the U.S. over the past
century. The pattern of abandonment of agriculture, reforestation followed by
urbanization has occurred in the PIE watershed as well (Schneider and Pontius
2001) (Fig 2). There have been
substantial changes in the frequency of storms along the U.S. Atlantic coast,
with the magnitude of change in storminess increasing at more northern
latitudes. At the PIE LTER changes in century-long storm frequency are
pronounced and precipitation and evapotranspiration have increased significantly
(Fig 3 Claessens et al. 2006). In the future, we can expect to see more
frequent, larger storms and higher storm frequency variability as well. There
have been decadal and century-long changes in sea level at PIE as well
(Fig 4). Therefore, the PIE LTER
study can be considered a microcosm for investigating the effects of these
globally important changes and it should thus be possible to transfer lessons
learned from the PIE LTER to
other coastal regions. 
There are numerous ways that these globally
important issues are affecting the biosphere. In the PIE LTER we focus on how these
issues influence organic matter and inorganic nutrient biogeochemistry and
estuarine foodwebs. The inputs of
organic matter and nutrients from land, ocean and marshes interact with the
external drivers (climate, land use, river discharge, sea level) to dictate the
extent and degree of nutrient and organic matter processing and determine the
spatial patterns of estuarine productivity and trophic structure. Our overarching question
is:
Within the context of this overarching question, our program addresses two hypotheses about the effects of driver variability and long-term change on ecosystem dynamics and the effects of inorganic vs. organic matter inputs on estuarine foodweb structure.
Hypothesis 1. The variability in land,
ocean and atmospheric forcing is a major factor controlling the fate of
allochthonous and autochthonous materials and the location and magnitude of
primary and secondary production.
All components of an estuarine ecosystem exhibit variation in space and time in response to external drivers (e.g., climate, river discharge and sea level) or in response to internal, biogeochemical and biological processes (e.g., nutrient cycling, population cycles and fish migrations). By comparing time-scale and intensity of processes, the relative importance of various physical or biological factors in controlling variation in process rates can be assessed and limits placed on the ability of one process to affect another. Processes that occur at similar scales are those likely to interact. Our research on the effect of changes in organic matter, nutrient and water fluxes on estuarine trophic structure and production requires that we evaluate variability in land and ocean drivers and assess the spatial and temporal scales over which the effects of these changes are likely to operate. Understanding the response of these systems to long-term changes in climate, sea level and land use first requires that we understand and can distinguish short-term variation from long-term trends.
Hypothesis 2. The interaction of inorganic nutrients with the quantity and quality of organic carbon and organic nitrogen plays an important role in determining the trophic structure, production and efficiency of estuarine food webs.
The delivery of organic matter and
nutrients has a major effect on estuarine food webs. We know from metabolic
studies that estuaries as a class are heterotrophic, consuming more organic
matter than produced (Hopkinson and Smith 2005) and thus dependent on
allochthonous sources. This is not
surprising as typically 1-2% of terrestrial NPP is exported to estuaries.
Most estuarine respiration is by bacteria (Hopkinson et al. 1989, Smith and Kemp
2003) due to the high C/N ratio and low lability of riverine organic matter (OM)
(Hopkinson et al. 1998). Elevated inputs of inorganic N from watersheds have
perhaps the greatest impact on estuarine condition. N-enrichment
(eutrophication) causes shifts in algal abundance, increases in algal
productivity and standing stocks, and in extreme cases, hypoxia, anoxia and fish
kills (Driscoll et al. 2003a, 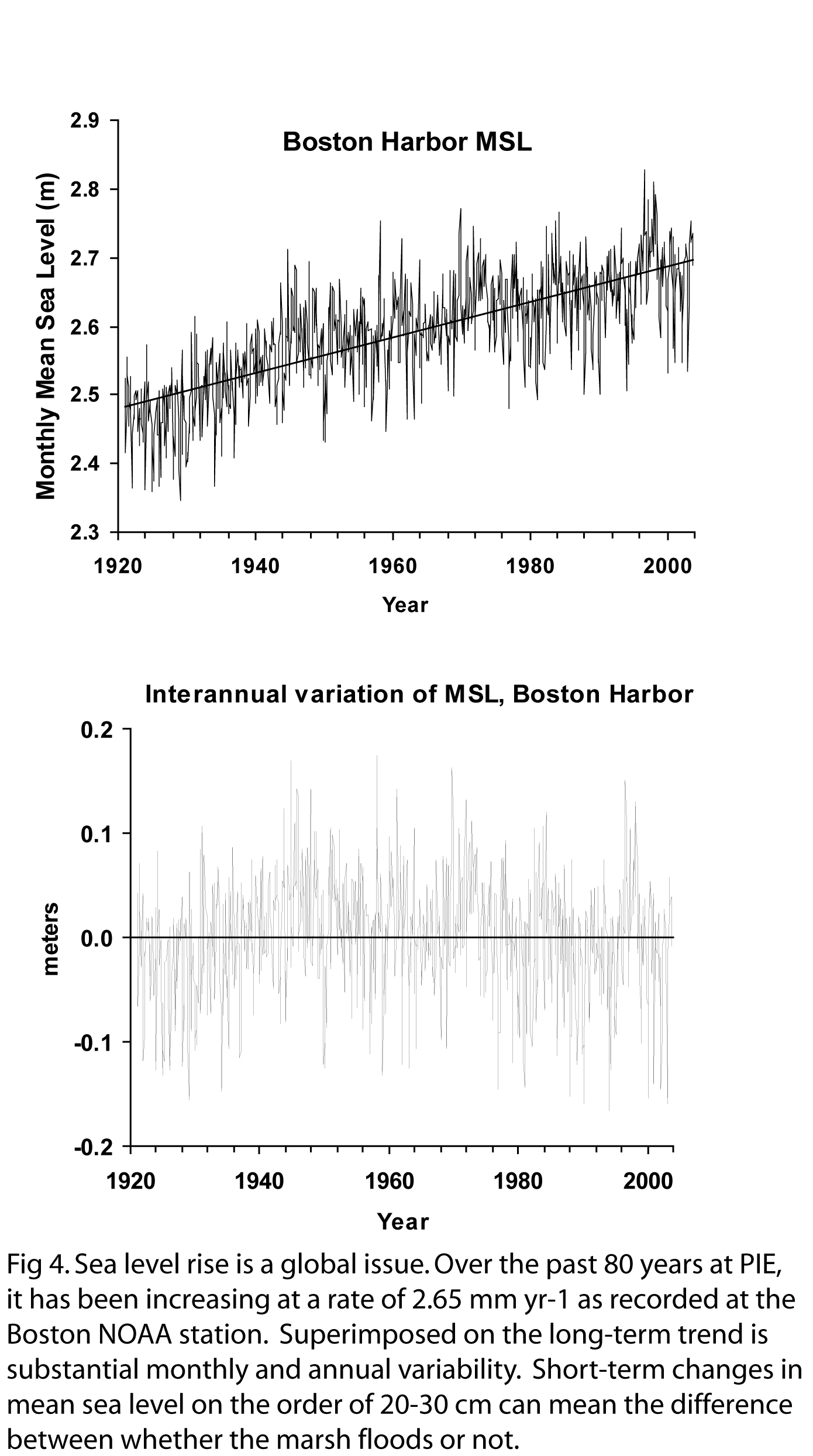 b). As watersheds become developed, organic
matter export declines and inorganic nutrient export increases (Raymond and
Hopkinson 2003, Pellerin et al. 2004, Wollheim et al. 2005, Williams et al.
2004, 2005). Major factors
controlling the autotrophic-heterotrophic balance of estuaries include the
balance between inputs of inorganic and organic nutrients (Hopkinson and Smith,
in press), water residence time and the lability of allochthonous organic matter
(Hopkinson and Vallino 1995). With
increased N enrichment, decreased organic matter inputs and perhaps decreased
freshwater inputs to estuaries in the future, we can expect changes in foodweb
structure, shifting from organic matter, microbially-based to inorganic
nutrient, phytoplankton-based (Deegan et al. 1994).
b). As watersheds become developed, organic
matter export declines and inorganic nutrient export increases (Raymond and
Hopkinson 2003, Pellerin et al. 2004, Wollheim et al. 2005, Williams et al.
2004, 2005). Major factors
controlling the autotrophic-heterotrophic balance of estuaries include the
balance between inputs of inorganic and organic nutrients (Hopkinson and Smith,
in press), water residence time and the lability of allochthonous organic matter
(Hopkinson and Vallino 1995). With
increased N enrichment, decreased organic matter inputs and perhaps decreased
freshwater inputs to estuaries in the future, we can expect changes in foodweb
structure, shifting from organic matter, microbially-based to inorganic
nutrient, phytoplankton-based (Deegan et al. 1994).
Early on we developed a trophic flow model and diagram (Fig 5) to synthesize our understanding of the effects of organic matter and nutrient inputs on trophic structure and function (Deegan et al. 1994). We envisioned 4 basic pathways of organic matter and nutrient processing that varied over decadal time scales in relation to long-term changes in the loading of organic matter and nutrients from watersheds:
1) the
classical grazing food chain: inorganic nutrients - phytoplankton > 20 µm -
macrozooplankton - planktivores - piscivores, 2) the microbial loop: organic matter
- bacteria - microflagellates - microzooplankton - macrozooplankton -
planktivores - piscivores, 3) a hybrid of the grazing food chain
and the microbial loop: inorganic nutrients - phytoplankton < 20 µm -
microflagellates - microzooplankton (the microbial loop) - macrozooplankton -
planktivores - piscivores, and 4) a
benthic food chain: organic matter - to sediments - bacteria - benthic meio
- and macrofauna - benthivores - piscivores.
This conceptualization, while
perhaps a good representation of long-term equilibrium conditions, over
emphasized the importance of organic matter and nutrient inputs from the
watershed and does not reflect the dynamic variability of estuaries. Because
riverine organic N inputs are greater than inorganic N inputs (Williams et al.
2004, 2005), our flow analysis model suggested a hybrid grazing -microbial loop
pathway dominating in the upper estuary with keen competition for nutrients
between phytoplankton and bacteria.
We expected to find evidence of terrestrial organic C fueling higher
trophic levels. We hypothesized
that the benthic food chain would take on importance down estuary, where
piscivore production would be supported from low quality organic matter inputs
from the marsh. Our temporal focus
was primarily decadal, the time scale over which we expected to see a shift
toward the classical food chain as urbanization increased DIN loading relative
to organic N.
Our perceptions of
how estuarine ecosystems are structured and function have changed substantially
over the past 6 years. We find that
the effect of watershed OM or DIN inputs is greatly modulated by river
freshwater discharge (Deegan and Garritt 1997, Holmes et al. 2000). We find that the most important input of
organic matter is the marsh (Raymond and Hopkinson 2003) and that there are
major recycling sources of DIN internal to the system that are controlled by
salinity variations (Giblin et al. submitted). We have yet to observe the classical
pelagic grazing food chain, rather phytoplankton blooms appear linked to higher
trophic levels equally through both pelagic and benthic webs (Fig 6; Hughes
et al. 2000). Benthic microalgal production is often an important base of the
foodweb (Fig
6; Tobias et al. 2003a, b). 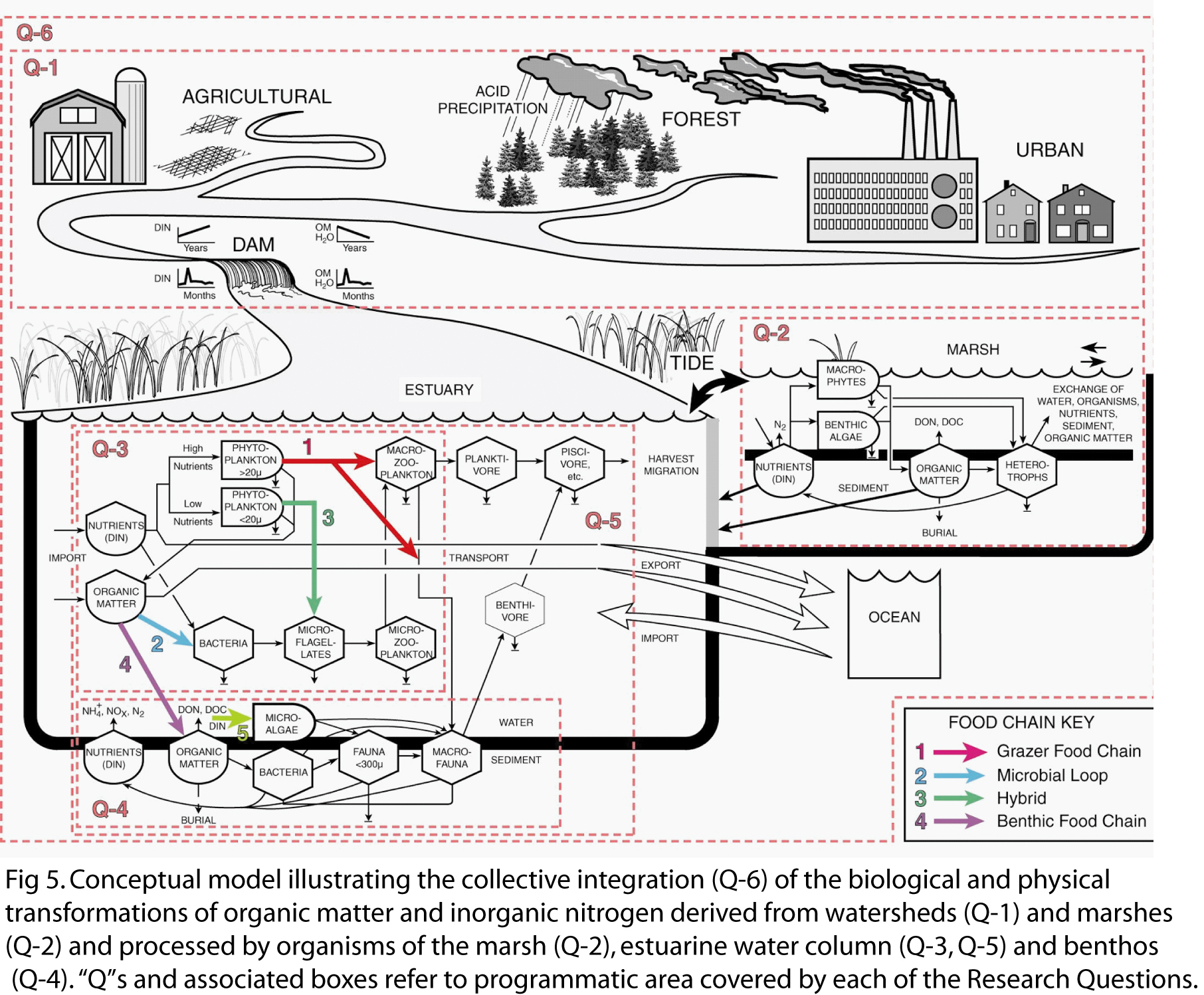
Freshwater discharge is of paramount importance in controlling foodweb structure because of its influence on residence time of water throughout the estuary (Vallino and Hopkinson 1998). Interestingly, only Parker River flow is an important factor. Although Ipswich River flow is 10x the Parker, its input at the mouth of the estuary has no discernible effect on residence time or salinity within Plum Island Sound (Vallino and Hopkinson 1998). Residence time of water in the upper estuary decreases from 34 2 d as river flow increases from 0.01 to 10 m3 s-1 (Vallino and Hopkinson 1998). In the lower estuary tidal mixing has the greatest effect on residence time; seasonal increases in river flow only decrease residence time from 1.2 - 0.6 d. The relative importance of physical or biological factors depends on the time scales of physical and biological processes. The influence of organic matter and nutrient inputs on the estuarine food web is dictated by the time scales of important processes relative to water residence time. Thus, by controlling estuarine residence time, the interplay of freshwater discharge and tidal regime defines the template upon which ecological processes can operate. For example, algal blooms can only occur when water residence time is substantially longer than bloom doubling time. Hence algal blooms only occur during very low flow during summer (Holmes et al. 2000).
We also find that there are spheres of influence that vary over the length of the estuary and over seasonal or shorter time scales, where foodwebs develop in response to physical drivers plus DIN and OM inputs. We have modified the trophic flow diagram (Fig 5) to include benthic microalgae and a microalgal food chain, and to include a direct link between phytoplankton blooms and benthic macrofauna. Figure 7 illustrates the interplay between external drivers as they define a template upon which the effects of DIN and OM inputs on foodweb structure operate. The figure shows examples of 4 foodweb structures that develop under different hydrologic templates and loading conditions. The spheres of influence identify where these foodweb structures are likely to be found. In no sense do we mean to imply that foodwebs are static communities controlled exclusively from the bottom-up. Foodwebs are dynamic and feedbacks between organisms and environment are to be expected.
One result of our altered perceptions is that the role of watershed DIN and OM inputs must be assessed within the context of the hydrologic template. The vast majority of inputs occur during late winter / early spring, when biological activity is at its lowest. We need to determine the extent and the mechanisms by which any of these inputs are retained for use later in the year. Marshes are net sinks for C and N; what enables them to export as much C and N as it appears they do and still remain net sinks? We hope to resolve some of these uncertainties as our research program continues.
Research Questions To address our overarching research question and hypotheses, we organize our research around 5 interrelated questions that define Programmatic Areas. The scope of each question and the manner in which each fits in the overall program is illustrated with numbered boxes around portions of our conceptual diagram (Fig 5).
Q1. What is the magnitude and long-term pattern of freshwater runoff and organic carbon and nitrogen and inorganic N loading from watersheds to the estuary?
Q2. How are tidal marsh processes and their connections to estuarine waters controlled by changes in land, atmospheric and oceanic forcing?
Q3. How do planktonic community structure and production respond to short and long-term changes in watershed runoff and the inputs of organic matter and nutrients?
Q4. How do benthic recycling of nutrients and processing of organic matter respond to changes in freshwater runoff and the quality and quantity of organic matter inputs?
Q5. How do the structure and function of higher trophic levels respond to changes in land, atmospheric and oceanic forcing as well as fisheries harvest?

{kind=link}
{kind=link}