
PIE-LTER Summary of Research Results through 1997
Watershed Keynote-
Major upland NO3- loads, primarily from urban areas, are
retained with 80-90% efficiency in rivers resulting in DON being the major form
of N export to estuary.
Land use, economic activity and population are changing rapidly in the three river basins comprising the 585 km2 upland watershed of the Plum Island Sound Estuary. We have 1) characterized historical landuse for the Parker, Rowley and Ipswich River watersheds, 2) developed digital databases (soils, land cover, slope, streams, erosivity, etc.) for use in hydrologic modeling, 3) characterized runoff chemistry for several small catchments and 4) modeled water, organic matter and nutrient discharge to the estuary (Finn and Hopkinson, 1997).
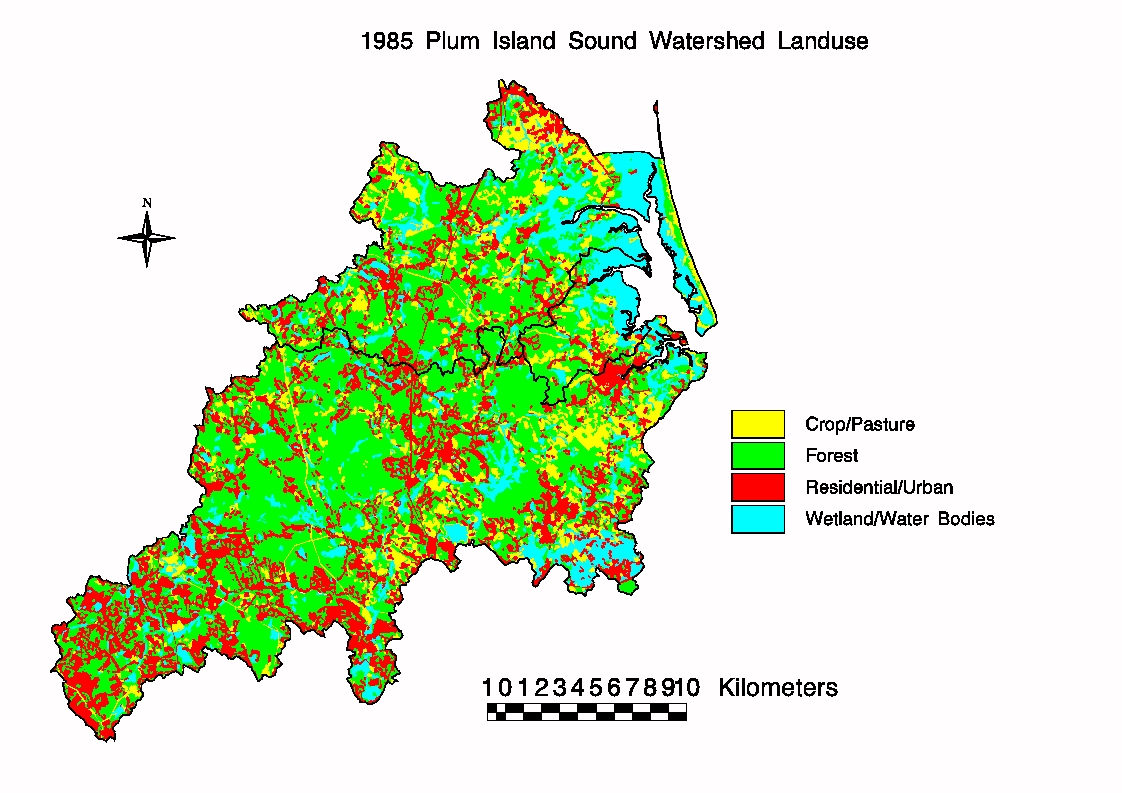 Land cover (Fig 1) in the watershed in 1985 was » 50% forest, 25% urban/suburban, 13% wetland,
and 12% agricultural. During the past 25 years urban area increased
substantially, largely at the expense of agricultural (down 50%) and forested
land (down 8%).
Land cover (Fig 1) in the watershed in 1985 was » 50% forest, 25% urban/suburban, 13% wetland,
and 12% agricultural. During the past 25 years urban area increased
substantially, largely at the expense of agricultural (down 50%) and forested
land (down 8%).
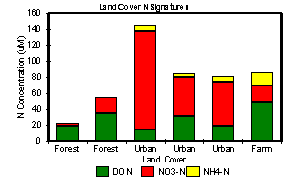
Runoff characteristics and streamwater chemistry
(spm, C, N and P) of streams draining various land covers differed
substantially. Total N was dominated by the dissolved fractions for all land
covers (Fig 2). Total dissolved N (TDN) concentration was highest in
urban runoff and least in forest runoff. While TDN was dominated by inorganic N
(primarily NO3-) in urban runoff, dissolved organic N
(DON) comprised up to 90% of the total in forested streams. Agricultural runoff
was intermediate in concentration, with inorganic and organic fractions of
roughly equal concentration.
Material loading and export budgets were created by scaling subcatchment
results to entire watersheds using a combination of hydrologic models (TR-55,
Brook6, and GWLF). Our riverine budgets did not balance. For N we estimated
515-975 Mg N yr-1 entered the river, while only 91-106 Mg yr-1
were discharged to the estuary. Thus in stream processing removed 80-90% of
inputs from land probably through a combination of processes including
vegetation uptake, sedimentation, organic matter accumulation and burial and
denitrification. DON was the major form of N exported from the watershed.
Estuarine Hydrodynamics Keynote- Residence time varies spatially and temporally from as long as 34 days in upper estuary to as little as 0.5 d in lower estuary depending on river flow.
Salinity distributions, dye release studies and parameter estimation techniques were used to determine longitudinal dispersion coefficients in the Estuary. Two hydrodynamic models were developed: 1) a 1-D tidally averaged, advection-dispersion model (Vallino and Hopkinson in press) and 2) a 1-D advection-dispersion model that accounts for marsh flooding (Vorosmarty and Loder 1994). These models were used to investigate characteristic mixing-time scales, average age, residence time and transit time, and to contrast nutrient dynamics during spring and neap tides. These time scales vary from hours to 2 months depending on location and freshwater discharge. Transport in the upper estuary can be either advection or dispersion dominated depending on discharge, while the lower estuary is always dominated by dispersive terms.
Microbial Utilization and Composition of Watershed-derived Organic Matter Keynotes - Novel bioenergetic model replaces Monod growth kinetics; Organic matter from various land covers differs in chemical characteristics, ability to support bacterial growth, and potential for N immobilization.
Effort focused on chemically characterizing organic matter from the variety of estuarine sources including plankton, macrophytes and runoff from several land covers. Bioassays were used to measure organic matter lability, patterns of nutrient immobilization or remineralization and bacterial growth rates and efficiency (Normann et al. 1995, Hullar et al. 1996). This work is the cornerstone of our research on organic matter - nutrient interaction effects on food web structure and efficiency.
Bioenergetic Model: A bioenergetic model was developed to examine growth dynamics associated with bacterial utilization of DOM, NH4+ and NO3- (Vallino et al. 1996). A novel, optimization approach was taken that provides more information on bacterial growth kinetics than Monod-type models that are typically employed to describe bacterial growth. The model provides a means to predict bacterial carbon yield, growth rate and N processing and is a bridge between chemical and biological characterization of DOM: bacterial dynamics are predicted from DOM chemical characteristics. Model results demonstrate that bacterial growth can not always be explained by a single constraint (such C:N ratio of substrate) as bacteria allocate resources to maximize growth rate subject to kinetic, thermodynamic and mass balance constraints. The model suggests that bacterial growth and yield can be predicted from the degree of reduction of the substrate (y ). We have been devoting considerable time applying the model to field situations and attempting to differentially characterize labile and recalcitrant pools of bulk DOM in natural waters.
Bioassays: Globally, changes in watershed
landuse have increased levels of organic carbon transported to the sea by 3-5
times over natural levels. Yet it is unclear how much of the DOM entering
estuaries is actually used by microbes and contributes to the food web and how
much resists degradation and is exported offshore. 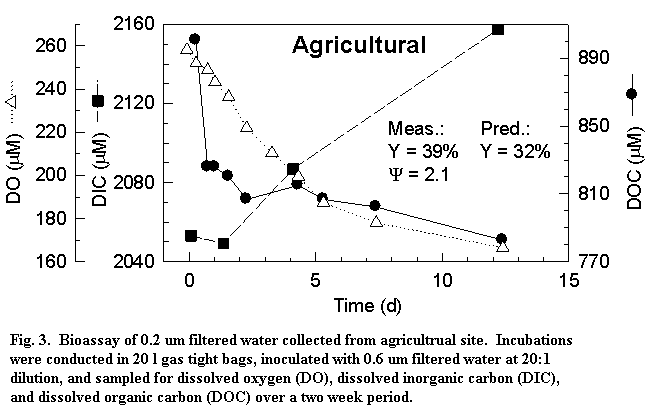 To better understand this, we have taken advantage of
"natural" differences in DOM composition and conducted experiments,
during high and low flow periods, with runoff water from urban, agricultural
and forested land covers (Uhlenhopp et al. 1995, Hobbie et al. 1996). DOM from
these sites ranged in concentration from 310 to 870 µM C, C/N from 10 to 40,
and DIN from <10 to > 50 µM N. Lability was measured with bacterial
growth bioassays where bacterial #s, DO, DIC, DOC, DON and nutrients were
monitored over time. Results (Fig 3) showed that labile DOC pools ranged
from ~40 µM C in the forest to ~100 µM C in the agricultural runoff and did not
vary much over the course of a year. The percent of the bulk pool that was
labile was small, similar between sites (5-15%) and varied little over time.
Bacterial yield during the first 3 d was compared to the bacterial yield
predicted by the bioenergetic model based on the estimated degree of reduction
of the DOM. Predicted bacterial yield (29, 32 and 43%) was in good agreement
with that observed (29, 39, 52%: Fig 3). We further observed that N was
remineralized during low flow when DIN was generally low and immobilized during
high flow when DIN was high.
To better understand this, we have taken advantage of
"natural" differences in DOM composition and conducted experiments,
during high and low flow periods, with runoff water from urban, agricultural
and forested land covers (Uhlenhopp et al. 1995, Hobbie et al. 1996). DOM from
these sites ranged in concentration from 310 to 870 µM C, C/N from 10 to 40,
and DIN from <10 to > 50 µM N. Lability was measured with bacterial
growth bioassays where bacterial #s, DO, DIC, DOC, DON and nutrients were
monitored over time. Results (Fig 3) showed that labile DOC pools ranged
from ~40 µM C in the forest to ~100 µM C in the agricultural runoff and did not
vary much over the course of a year. The percent of the bulk pool that was
labile was small, similar between sites (5-15%) and varied little over time.
Bacterial yield during the first 3 d was compared to the bacterial yield
predicted by the bioenergetic model based on the estimated degree of reduction
of the DOM. Predicted bacterial yield (29, 32 and 43%) was in good agreement
with that observed (29, 39, 52%: Fig 3). We further observed that N was
remineralized during low flow when DIN was generally low and immobilized during
high flow when DIN was high.
Organic Matter - Nutrient Interaction Effects on Higher Trophic Levels: Keynotes - Mesocosm experiments support trophic model analysis of the effect of organic matter on foodweb efficiency; stable isotope analysis of food sources and consumers indicates preferential utilization of low quality marsh detritus by benthivores.
Mesocosm Experiments: We conducted
experiments in 10 m3 plastic bags to examine differential effects of
phytoplankton derived organic C vs terrestrial organic C on planktonic
community structure and production. To 4 mesocosms with natural, intact
planktonic communities (through larval Menidia fish) were added either: A)
control - nothing, B) DOM from dead leaf litter leachate: final DOC up
from ~100 to 500 µM, C) daily additions of inorganic N, P and Si, and D)
leaf leachate DOM as in (B) plus daily additions of nutrients as in (C). DI13C
was added to each bag to trace phytoplankton derived C through the food web.
Dramatic differences in community structure and production resulted. Gross
production and net community production were highest in the nutrient enriched
(C) treatment and lowest in the litter leachate (B) treatment. Whereas primary
production was dominated by diatoms > 20 µm in the nutrient enriched bag,
85-90% of production was attributed to phytoplankton < 20 µm in the control
(A) and DOM (B) bags. Food web efficiency was low in the DOM amendment bags
(B&D) with 80% of respiration attributable to organisms < 1 µm and only
20% attributable to organisms > 20 µm. In contrast, trophic transfer was
substantially greater in the nutrient addition bag as evidenced by ~50% of
system respiration being attributable to organisms between 20-80 µM and >80
µm in size. Less than 50% of total respiration was attributable to bacteria.
Species composition changed with treatments as well. In the DOM bag (B), Podon
and Parvocalanus became the dominant zooplankton while in the DIN bag
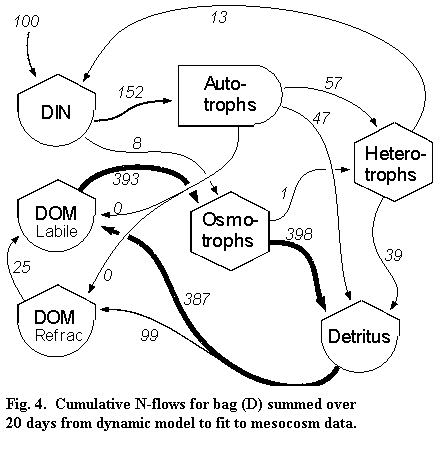
(C), Acartia quickly became dominant. A N-flow model for bag (D)
illustrates the dominance of the microbial loop and the trivial transfer of N
to higher trophic levels (Fig 4). These results confirm previous flow
model predictions about the effects of organic matter vs nutrients on trophic
structure and production (Deegan et al. 1996): inorganic nutrients support a
short, efficient and productive food web while low quality organic matter
without inorganic nutrients support an inefficient, microbe-dominated food web
with low levels of small phytoplankton production.
Sources of Organic Matter Supporting Higher Trophic Levels in the Estuary: The importance of phytoplankton, benthic microalgae, marsh grass and terrestrial organic matter to the estuarine food web was evaluated using stable isotopes of C, N and S (Deegan and Garritt 1997). Strong spatial differences in relative importance of organic matter sources existed within the estuary. While there is substantial spatial heterogeneity in organic matter sources within the estuary, consumers utilize sources produced in the same region. Terrestrial organic matter was not evident in the food webs of the middle and lower estuary and its importance in the oligohaline region was equivocal as there was not a distinct separation in isotopic signatures between fresh marsh grass and terrestrial organic matter. There was an interesting dichotomy in the relative importance of phytoplankton vs marsh detritus in the diets of pelagic vs benthic organisms. Benthic consumers reflected a greater dependence on marsh detritus.
Macrofaunal Community Structure Keynotes - Low number of fish species in estuary reflects nature of Acadian Biogeographic Province; Six-fold increase in forage fish density between 1965 and 93/4.
The current status of the estuary as a habitat for
important marine fish and shellfish was ascertained using seine and trawl
samplings (Buchsbaum 1996). Sampling sites and techniques were similar to those
of earlier surveys conducted by the Massachusetts Division of Marine Fisheries
(Jerome et al. 1967). The distribution of organisms varied in the different
habitats of the estuary. In open water and sandy substrata, Menidia, Fundulus
and Crangon dominated. In muddy salt marsh habitats the same species
were present but Fundulus was the dominant and Palaemonetes was
also common. In brackish, riverine habitats Morone, Alosa and Rhithropanopeus
were the most common organisms. In comparison to the 1965 survey, the average
density of fish was about 6-fold higher in 1993-94. In both studies Fundulus
and Menidia were dominants in the community but species richness based
on seine sampling was slightly higher in 1993-4. Explanations for the
differences in density between years is not obvious, but may be related to
marked differences in average sea level or river discharge during critical
periods in the development of these organisms. Long term data are needed to
adequately explain whether the differences between 1965 and 1993/4 are caused
by regional and environmental factors or whether fish populations naturally
exhibit short term variability.
Nutrient Cycling and Metabolism Keynotes - Temporal variability in tidal amplitude and river discharge affect spatial patterns of nutrient cycling, benthic nutrient fluxes and primary production; Estuarine waters are heterotrophic and therefore net sources of CO2 and inorganic nutrients.
Temporal and spatial aspects of metabolism and
nutrient cycling were documented, including primary production (Alderman et al.
1995, Balsis et al. 1995), spring-neap tidal contrasts in marsh-water organic
matter and nutrient exchange (Vorosmarty and Loder 1994), benthic metabolism
and nutrient exchange (Hopkinson et al. submitted), and inorganic nutrient and
dissolved organic matter dynamics. 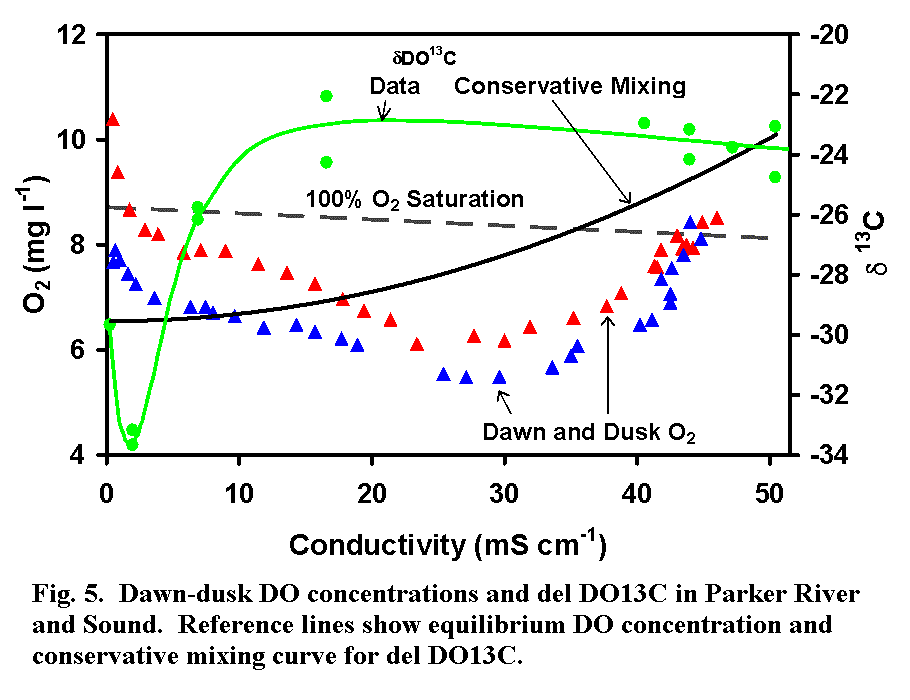 High primary production occurs in the lower estuary
during winter in association with the Gulf of Maine spring bloom. Production is
high during summer in the oligohaline region but only when mixing time scales
are long relative to phytoplankton turnover time. The benthos is a major site
of organic matter degradation and because of denitrification and the high C/N
nature of organic matter processed in the benthos, it exerts a net
heterotrophic influence on the system (ie, C production from recycled N
<< organic carbon remineralized). Seasonal and annual variations in
porewater salinity of oligohaline and brackish sediments alters the timing of
NH4+ release/storage in sediments. The overall system is
heterotrophic with DO often depressed to 60% of saturation in mid-estuary (Fig
5). The region of highest heterotrophy coincides with organic matter inputs
from marshes as evidenced by dDO13C
gradients in the water column (Fig. 5).
High primary production occurs in the lower estuary
during winter in association with the Gulf of Maine spring bloom. Production is
high during summer in the oligohaline region but only when mixing time scales
are long relative to phytoplankton turnover time. The benthos is a major site
of organic matter degradation and because of denitrification and the high C/N
nature of organic matter processed in the benthos, it exerts a net
heterotrophic influence on the system (ie, C production from recycled N
<< organic carbon remineralized). Seasonal and annual variations in
porewater salinity of oligohaline and brackish sediments alters the timing of
NH4+ release/storage in sediments. The overall system is
heterotrophic with DO often depressed to 60% of saturation in mid-estuary (Fig
5). The region of highest heterotrophy coincides with organic matter inputs
from marshes as evidenced by dDO13C
gradients in the water column (Fig. 5).
Whole System Synthesis
Keynote - Generalized estuarine metabolism
model evaluates patterns of production, nutrient cycling and
autotrophy/heterotrophy caused by variations in river discharge and material
loading from land.
Flow model analysis and simulation models have been used from the outset of our
research program to guide research, help in designing experiments, synthesize
results of microcosm and mesocosm experiments and to extend experimental and
process based research results to the field. In addition to models described
above, we have used flow model analysis to understand the effects of organic
matter - nutrient interactions on estuarine trophic dynamics (Deegan et al.
1995). We developed a generalized estuarine metabolism model to explore the
effects of land use change and river modifications on patterns of production,
respiration and nutrient cycling (Hopkinson and Vallino 1995). The model was
useful for evaluating the effect of residence time, organic matter quality, and
DON:DIN and POM:DOM loading ratios on patterns of net ecosystem metabolism.
![]()
